


L-глутаматът е един от основните възбуждащи невротрансмитери в човешкия мозък и играе съществена роля в практически всички дейности на нервната система. В следващата статия ще обсъдим общите принципи на L-глутаматната сигнализация в мозъка. След това ще демонстрираме тази схема, като опишем различните групи от извънклетъчен глутамат, включително синаптичен, перисинаптичен и екстрасинаптичен, произтичащ от везикуларни и невезикуларни източници или анормално разположени глутаматни рецептори извън синапсите, както и да обсъдим техните възможни физиологични функции в човешкия мозък.
Съдържание
Глутаматна сигнализация в мозъка
Според изследвания, човешкият мозък има около 6 до 7 ?mol/g мокро тегло на L-глутамат. L-глутаматът, заедно с глутамин, е една от най-разпространените свободни аминокиселини в централната нервна система (ЦНС). Преди повече от пет десетилетия няколко изследователски проучвания показаха, че L-глутаматът има възбуждащ отговор върху нервните клетки. Оттогава ролята му като възбуждащ невротрансмитер, както и неговият мозъчен метаболизъм са оценени в множество научни изследвания.
L-глутаматът обикновено се намира в синаптичните везикули в пресинаптичния терминал чрез процеса на везикуларни глутаматни транспортери. Освен това, няколко от L-глутамата във везикулите могат да се развият от аспартат аминотрансфераза, свързана с везикула от 2-оксоглутарат, използвайки L-аспартат като донор на аминогрупата. По време на деполяризацията на пресинаптичната мембрана, L-глутаматът се освобождава в синаптичната цепнатина и се свързва с йонотропни глутаматни рецептори, известни като iGluRs, в постсинаптичната мембрана, както е показано на Фигура 1. Според изследователски проучвания, iGluRs се характеризират като лиганд- затворени йонни канали, които включват рецептори от типа а-амино-3-хидрокси-5-метил-4-изоксазол пропионова киселина (АМРА), каинат и N-метил-D-аспарагинова киселина (NMDA). Докато AMPA и каинатните рецептори основно регулират и поддържат притока на натрий, NMDA рецепторите всъщност имат висока калциева проводимост. Освен това, активирането на NMDA рецепторите играе основна роля в синаптичната пластичност и ученето. За разлика от другите iGluRs, активността на NMDA рецепторите в крайна сметка е ограничена от Mg+2 блок при нормалния мембранен потенциал, но йонният канал незабавно се отблокира чрез деполяризация на мембраната, която елиминира Mg+2 от порите. Освен това, NMDA рецепторите са тетрамери, които имат две NR1 субединици и две NR2 или NR3 субединици, според няколко изследователски проучвания.
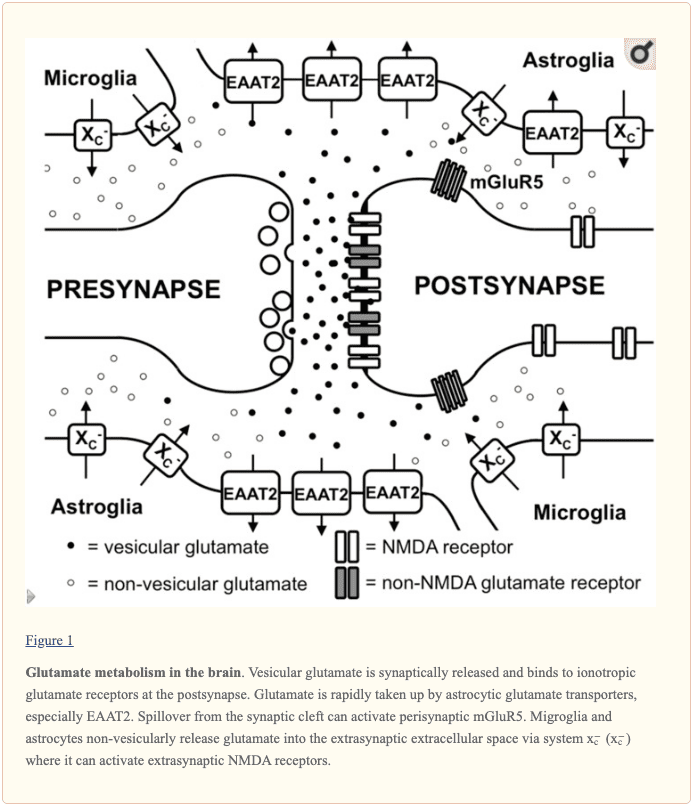
В допълнение към iGluRs, има и осем изоформи на метаботропни глутаматни рецептори (mGluRs), които принадлежат към семейството на G-протеин-свързани рецептори, където те не развиват йонни канали, а вместо това сигнализират чрез различни системи за втори месинджър. Деполяризацията, свързана с L-глутамат, причинява постсинаптичен възбуждащ потенциал, който улеснява развитието на потенциал за действие в хълма на аксона. Глутаматергичният синапс се активира от астроцитни процеси, които демонстрират високи нива на възбуждащи аминокиселинни транспортери (EAATs). Има пет различни EAAT, EAAT1 до 5, от които EAAT1 и 2 са първичните астроцитни EAAT, докато EAAT3 показва предимно невронална експресия. Приблизително 90 процента от транспорта на L-глутамат се регулира и поддържа от EAAT2, като GLT-1 в моделите на гризачи. Тези транспортери след това съвместно транспортират 2 или 3 молекули Na+ и един протон с всяка молекула L-глутамат или L-аспартат заедно с обратния транспорт на K+ йон. Следователно, като използват електрохимичния градиент на тези йони в плазмената мембрана като енергиен източник, транспортерите са в състояние безопасно и ефективно да натрупват L-глутамат и L-аспартат в клетките срещу техните внезапни градиенти на концентрация между вътре и извън клетките. Това позволява на мозъка да контролира много ниска извънклетъчна концентрация на L-глутамат в ниския микромоларен диапазон. Обикновено се смята, че L-глутаматът, поет от астроцитите, се превръща в глутамин от ензима глутамин синтетаза, след това глутаминът се освобождава, поема се от невроните и се превръща в L-глутамат, където в крайна сметка се използва отново за невротрансмисия.
Екстрасинаптичен глутамат в мозъка
Освен основната роля на L-глутамата като първичен възбуждащ невротрансмитер, освободен от глутаматергичните пресинапси, както беше споменато по-горе, стана очевидно, че L-глутаматните рецептори извън синаптичната цепнатина също играят съществена роля във физиологията на мозъка. В малкия мозък беше демонстрирано чрез оценка на AMPA рецептор-медиираните течения в глията на Бергман, че концентрациите на синаптично освободен L-глутамат могат да достигнат екстрасинаптични концентрации до 190 цМ, докато концентрациите в синаптичната цепнатина могат да надвишават 1 mM. Освен това е показано, че няколко mGluRs демонстрират различна локализация в близост до постсинаптичната плътност, което би им позволило незабавно да разпознаят L-глутамат, излизащ от синаптичната цепнатина, както е показано на Фигура 1. Въпреки това, настоящите изследвания показват, че iGluRs, особено от типа NMDA, се намират и в екстрасинаптичните участъци в невронната клетъчна мембрана. Използвайки светлинна и електронна микроскопия, други изследователски проучвания също показват, че екстрасинаптичните NMDA рецептори се събират в различни области на близък контакт в дендритния вал с аксони, аксонни терминали или астроцитни процеси. Делът на екстрасинаптичните NMDA рецептори се оценява на 36 процента от дендритния NMDA рецепторен пул в срезовете на хипокампа на плъхове. Въпреки че екстрасинаптичните NMDA рецептори са свързани с подобни скелетни протеини като синаптичните NMDA рецептори, проучване in vitro предполага, че екстрасинаптичните и синаптичните NMDA рецептори могат в крайна сметка да активират различни сигнални пътища надолу по веригата с различни резултати, включително потискане на активността на CREB от екстрасинаптичен NMDA рецептор активиране, както и активиране от синаптични NMDA рецептори. Освен това, NMDA рецепторите, локализирани екстрасинаптично върху дендритните валове, свързват екстрасинаптичния L-глутамат, както и регулират и поддържат притока на Ca2+ по време на елиминирането на Mg+2 блока чрез деполяризация на дендрита по време на обратното действие на потенциалите на действие. Изследванията показват, че освобождаването на L-глутамат от астроцитите може да активира бавни вътрешни потоци през екстрасинаптични NMDAR рецептори в CA1 неврони, които също могат да бъдат в крайна сметка синхронизирани. Механизмите, чрез които глиалните клетки освобождават L-глутамат, както и как се контролират екстрасинаптичните концентрации на L-глутамат са жизненоважни за разбирането как се контролира активността на екстрасинаптичните NMDA рецептори.
Предложени са различни механизми, чрез които астроцитите могат да отделят L-глутамат, включително везикуларно освобождаване на L-глутамат и невезикуларно освобождаване през анионни канали, както и конексинови хемиканали и освобождаване през цистин/глутаматната антипортерна система x?c. Няколко изследователски проучвания категорично предполагат, че везикуларното освобождаване от астроцитите играе второстепенна роля, тъй като свързаното с Ca+2 освобождаване на L-глутамат все още присъства в астроцитите, създадени от доминантно-отрицателни SNARE мишки, където везикуларното освобождаване може да бъде блокирано чрез изтегляне на доксициклин. Системата x?c е цистин/глутамат антипортер, който се характеризира като хетеродимерни аминокиселинни транспортери, съставени от xCT като специфична субединица и 4F2hc като безразборна тежка верига. Този транспортер е демонстриран в мозъка, особено в астроглиални и микроглиални клетки, както е показано на Фигура 1. Фактът, че екстрасинаптичните нива на L-глутамат в различни региони на човешкия мозък са понижени с приблизително 60 процента до 70 процента при xCT нокаут мишки Изследователските проучвания показват, че системата x?c освобождава L-глутамат в екстрасинаптичното пространство и предполага, че този транспортер е от съществено значение за регулирането на екстрасинаптичните нива на L-глутамат. Това се подкрепя допълнително от наблюдението, че когато се измерва чрез микродиализа in vivo, увеличаването на екстрасинаптичния L-глутамат, разработен от EAAT инхибиторите, се неутрализира чрез блокиране на системата x?c, докато блокирането на освобождаването на невронния везикуларен L-глутамат е неефективно. Все още са необходими допълнителни изследвания.
Взети заедно, глутаматергичните невротрансмисии не се случват просто чрез класическите възбуждащи синапси, но също и чрез екстрасинаптичните L-глутаматни рецептори, както е показано на Фигура 1. И накрая, нивата на екстрасинаптичен L-глутамат се определят, поне частично, от глиални невезикуларни Освобождаването на L-глутамат, както е показано също на Фигура 1. Въпреки това, регулирането на екстрасинаптичните нива на L-глутамат, както и неговата времево-пространствена динамика и ефектът му върху невроналната функция, невродегенерация и поведение, далеч не са напълно разбрани от изследователи, здравни специалисти и пациенти.

Глутаматът, заедно с аспартата, е един от основните възбуждащи невротрансмитери в човешкия мозък. Въпреки че играе основна роля в цялостната структура и функция на нервната система, прекомерните количества глутамат могат в крайна сметка да причинят ексайтотоксичност, която може да доведе до различни здравословни проблеми, като болестта на Алцхаймер и други видове неврологични заболявания. Следващата статия описва ролята на глутамата в човешкия мозък. – Д-р Алекс Хименес DC, CCST Insight
Лечение на невропатия с LLLT
L-глутаматът е един от основните възбуждащи невротрансмитери в човешкия мозък и играе съществена роля в практически всички дейности на нервната система. В статията по-горе обсъдихме общите принципи на сигнализирането на L-глутамат в мозъка. След това демонстрирахме тази схема, като описахме различните групи от извънклетъчен глутамат, включително синаптичен, перисинаптичен и екстрасинаптичен, произтичащ от везикуларни и невезикуларни източници или ненормално разположени глутаматни рецептори извън синапсите, както и обсъдихме техните възможни физиологични функции в човешкия мозък. Обхватът на нашата информация е ограничен до хиропрактика, мускулно-скелетни и нервни проблеми, както и статии, теми и дискусии по функционална медицина. Ние използваме функционални здравни протоколи за лечение на наранявания или хронични заболявания на опорно-двигателния апарат. За допълнително обсъждане на темата по-горе, моля не се колебайте да попитате д-р Алекс Хименес или да се свържете с нас на 915-850-0900 .
Подготвен от д-р Алекс Хименес
Препратки
- Леверенц, Ян и Памела Махер. „Хронична глутаматна токсичност при невродегенеративни заболявания – какви са доказателствата?“ Граници в невронауката, Frontiers Media SA, 16 декември. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4679930/.
Допълнителна дискусия по темата: Хронична болка
Внезапната болка е естествен отговор на нервната система, който помага да се демонстрира възможно нараняване. Например, сигналите за болка пътуват от ранен регион през нервите и гръбначния мозък до мозъка. Болката като цяло е по-малко силна, тъй като нараняването наранява, но хроничната болка е различна от средната тип болка. При хронична болка човешкото тяло ще продължи да изпраща сигнали за болка до мозъка, независимо дали нараняването е заздравяло. Хроничната болка може да продължи от няколко седмици до дори няколко години. Хроничната болка може да повлияе изключително много на мобилността на пациента и може да намали гъвкавостта, силата и издръжливостта.
Неврален Zoomer Plus за неврологични заболявания

Д-р Алекс Хименес използва серия от тестове, за да помогне за оценка на неврологичните заболявания. Невронният зумерTM Плюс това е масив от неврологични автоантитела, който предлага специфично разпознаване на антитяло към антиген. Жизненият неврален зумерTM Плюсът е предназначен за оценка на реактивността на индивида към неврологични антигени 48 с връзки към различни неврологично свързани заболявания. Жизненият неврален зумерTM Плюсът цели намаляване на неврологичните състояния чрез овластяване на пациенти и лекари с жизненоважен ресурс за ранно откриване на риска и засилен фокус върху персонализирана първична профилактика.
Формули за поддръжка на метилиране

XYMOGEN на Ексклузивни професионални формули са достъпни чрез избрани лицензирани професионалисти в областта на здравеопазването. Продажбата по интернет и дисконтирането на формулите XYMOGEN са строго забранени.
Гордост, Д-р Александър Хименес прави формулите XYMOGEN достъпни само за пациенти под нашата грижа.
Моля, обадете се в нашия офис, за да можем да назначим консултация с лекар за незабавен достъп.
Ако сте пациент Медицинска и хиропрактична клиника за нараняване, можете да попитате за XYMOGEN, като се обадите 915-850-0900.
![]()
За ваше удобство и преглед на XYMOGEN продукти, моля прегледайте следната връзка. *XYMOGEN-подреждане иИзтегли
* Всички горепосочени политики на XYMOGEN остават в сила.
Отказ от отговорност
Професионален обхват на практика *
Информацията тук на "Функционална неврология: ролята на глутамата в мозъка" няма за цел да замени личната връзка с квалифициран здравен специалист или лицензиран лекар и не е медицински съвет. Насърчаваме ви да вземате здравни решения въз основа на вашите изследвания и партньорство с квалифициран здравен специалист.
Информация за блога и дискусии за обхват
Нашият информационен обхват е ограничено до хиропрактика, мускулно-скелетна, акупунктура, физикални лекарства, уелнес, допринасящи етиологични висцерозоматични нарушения в рамките на клинични презентации, свързана соматовисцерална рефлексна клинична динамика, сублуксационни комплекси, чувствителни здравни проблеми и/или статии, теми и дискусии от функционална медицина.
Ние предоставяме и представяме клинично сътрудничество със специалисти от различни дисциплини. Всеки специалист се ръководи от своя професионален обхват на практика и своята юрисдикция за лицензиране. Използваме протоколи за функционално здраве и уелнес за лечение и поддържане на грижи за наранявания или нарушения на мускулно-скелетната система.
Нашите видеоклипове, публикации, теми, теми и прозрения обхващат клинични въпроси, проблеми и теми, които се отнасят до и пряко или косвено подкрепят нашия клиничен обхват на практика.*
Нашият офис разумно се е опитал да предостави подкрепящи цитати и е идентифицирал съответните изследвания, подкрепящи нашите публикации. Ние предоставяме копия на подкрепящи научни изследвания, достъпни за регулаторните съвети и обществеността при поискване.
Разбираме, че обхващаме въпроси, които изискват допълнително обяснение как може да подпомогне определен план за грижи или протокол за лечение; следователно, за да обсъдите допълнително темата по-горе, моля не се колебайте да попитате Д-р Алекс Хименес, окръг Колумбия, Или се свържете с нас на адрес 915-850-0900.
Ние сме тук, за да помогнем на вас и вашето семейство.
Благословения
Д-р Алекс Хименес н.е., MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
имейл: coach@elpasofunctionalmedicine.com
Лицензиран като доктор по хиропрактика (DC) в Тексас & Ню Мексико*
Тексас DC Лиценз # TX5807, Ню Мексико DC Лиценз # NM-DC2182
Лицензирана като регистрирана медицинска сестра (RN*) в Флорида
Флорида Лиценз RN Лиценз # RN9617241 (Контролен номер 3558029)
Компактен статус: Многодържавен лиценз: Упълномощен да практикува в 40 състояния*
Понастоящем записан: ICHS: MSN* FNP (Програма за практикуваща семейна медицинска сестра)
Д-р Алекс Хименес DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Моята цифрова визитка
 Отново Ви приветстваме¸
Отново Ви приветстваме¸
